God and Nature Spring 2023
|

|
By Larry Funck
The primary question raised by Sy Garte in his article on assembly theory in the Winter 2023 issue of God and Nature involves its relevance to the question of the origin of life. Garte’s article reflects on the results of a recent paper in Nature about using an index of molecular complexity (which the authors called the Molecular Assembly, or MA, index) as a way to detect life in distant worlds. I was curious if there were other comments in the literature regarding possible implications of assembly theory for the origin-of-life question, so I checked Web of Science to find citations for this Nature paper. I have discovered that essentially all of them were in articles related to exploratory astrobiology.
The one exception was a “Perspective” article in Life entitled “The Grayness of the Origin of Life” by Smith, Hillary H. et al. (1). The emphasis in this article is on the severe challenges that arise from the numerous unknowns inherent in the origin of life. Among these is the definition of the phenomenon itself. What is life? Is there a threshold that would provide criteria for distinguishing between a system classified as abiotic and one classified as biotic? The authors’ answer is in the negative. Instead, they suggest a gradual progression between the non-living and the living resulting from the “protracted evolutionary process that gave rise to life” (2). They describe the progression from abiotic to biotic, or geochemistry to biochemistry, as involving a spectrum over time with no clear distinctions, but rather a lot of “grayness.”
The primary question raised by Sy Garte in his article on assembly theory in the Winter 2023 issue of God and Nature involves its relevance to the question of the origin of life. Garte’s article reflects on the results of a recent paper in Nature about using an index of molecular complexity (which the authors called the Molecular Assembly, or MA, index) as a way to detect life in distant worlds. I was curious if there were other comments in the literature regarding possible implications of assembly theory for the origin-of-life question, so I checked Web of Science to find citations for this Nature paper. I have discovered that essentially all of them were in articles related to exploratory astrobiology.
The one exception was a “Perspective” article in Life entitled “The Grayness of the Origin of Life” by Smith, Hillary H. et al. (1). The emphasis in this article is on the severe challenges that arise from the numerous unknowns inherent in the origin of life. Among these is the definition of the phenomenon itself. What is life? Is there a threshold that would provide criteria for distinguishing between a system classified as abiotic and one classified as biotic? The authors’ answer is in the negative. Instead, they suggest a gradual progression between the non-living and the living resulting from the “protracted evolutionary process that gave rise to life” (2). They describe the progression from abiotic to biotic, or geochemistry to biochemistry, as involving a spectrum over time with no clear distinctions, but rather a lot of “grayness.”
The high MA values of biotic molecules may be interpreted as another indication of how extraordinarily life-friendly this Creation is. |
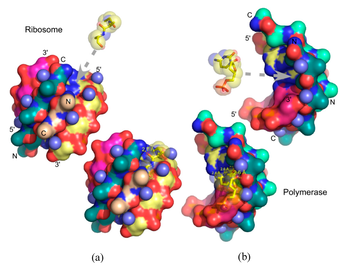
Smith et al.’s approach draws upon the “chemoton” model of Gánti to provide a minimal model of life, comprising the three fundamental subsystem functions—compartmentalization, metabolism, and an information system for reproduction (3). Interestingly, their reference to the Nature paper occurs in the context of a discussion about the requirements for information storage via RNA, where they make the point that “the ability to store useful information is determined by the length of the encoding string coupled with maintenance of non-random sequences selected by evolution over time” (4). They relate assembly theory to the origin-of-life question in the context of the origin of information storage and transmission, but do not draw any direct conclusions from the theory.
In my opinion the critical juncture between assembly theory and origin of life involves the fundamental question of the probability of life’s occurrence. As Garte suggests in his article, molecules with a higher molecular assembly (MA) index, especially the critically important nucleotides and their polymers, molecules involved in information storage and transmission, would be hard to produce abiotically.
I would suppose that assembly theory proponents would argue that there is an inverse correspondence between a molecule’s MA value and the probability of that molecule occurring in the absence of life. I suspect that they would want to leave open the question of the probability of the origin of life itself.
If the production of high MA molecules needed for life’s origin had to occur in advance of its origin, then it would follow that life’s spontaneous appearance would have an exceedingly low probability. But what if life emerged through a “protracted evolutionary process” that also produced the high MA molecules as part of that process? Then both living systems and high MA molecules associated with life would be the outcome of life’s origin. The question boils down to this: What were the chemical pathways to the production of systems possessing high MA molecules—systems capable of exhibiting life-like behavior, and therefore classifiable as biotic—or, in simpler terms, how did life begin?
Origin-of-life theoretician Stuart Kauffman wrote in 1995 (still true today): “Anyone who tells you that he or she knows how life started on the sere earth… is a fool or a knave” (5).
Origin-of-life theories are abundant and varied. For example, there are the geneticists versus the metabolists. Which had primacy—the production of energy/metabolites or the origin of information coding and reproductive capability? What constituted the first forms of life, and what did they eat—were they heterotrophic, incorporating organic substances, or were they autotrophs, making their own organics from inorganic starting materials in their initial formation and their continued maintenance?
One of the suggested ways that a geochemical to biochemical transition is imagined involves the proposal of catalytic activity by metal atoms/ions on mineral surfaces that initiated the first versions of metabolic processes and led to the development of metalloenzymes. There are plenty of examples of speculative theory along these lines, most extensively in the proposals for life’s origin at deep sea hydrothermal vents.
Perhaps the best experimental support of an origin via autotrophic metabolism comes from recent discoveries of non-enzymatic metabolism. Markus Ralser details in his 2018 review article (6) multiple examples discovered within the last decade of entire metabolic pathways catalyzed by metal ions, particularly Fe(II). He argues that this provides substantial support for the idea that biochemical pathways originated in an abiotic, geochemical context. Ralser suggests that by the use of appropriate analytical techniques, it is likely that many more examples of non-enzymatic metabolic pathways will be discovered.
Muchowska, et al., in a more recent review article (7), discuss the concepts advanced by Ralser and extend the consideration to the possibility of non-enzymatic synthesis pathways for nucleotides that would parallel their biosynthetic pathways. They suggest that the lack of current evidence for such pathways is likely a consequence of the lack of research along those lines. They also argue in favor of a “systems” approach to prebiotic chemistry, which would investigate the possibility of cooperative development of nucleic acids and polypeptides.
So how does this relate to the question of high MA molecules before life’s emergence? In short, the occurrence of complex metabolic pathways driven not by high MA enzymes but by very low MA metal ions suggests the possibility that the high MA molecules may have emerged in the process of the gradual progression towards systems that would have increasingly biotic characteristics and be eventually characterized as alive.
Arguably the most critical step in the progression towards life involved the appearance of the genetic coding system and accompanying enzymatic apparatus. There are several theories about how this occurred, but none has gained wide acceptance to date. In my opinion, the most attractive one has been advanced by Carter and Wills (8). Their theory, which is empirically based, rests principally on a proposal of intimate cooperation between RNA and peptides in the emergence of the code through the formation of “complexes involving structurally complementary oligonucleotides and dipeptides” (9). These complexes are proposed to have originated long before genuine code expression occurred involving simple versions of tRNA and aaRS proteins.
My guess is that were Carter or Wills to be asked about the existence of these relatively high MA value molecular complexes well in advance of a functional coding system, they would probably reply in a manner similar to that proposed above—the high MA oligonucleotide/dipeptide complexes were merely intermediates on the path to life in the “protracted evolutionary process.”
So what should we conclude? Should the high MA values for biomolecules be viewed as problematic for the origin-of-life theoretician? I would suggest they will be troublesome if the theoretician is uncomfortable living in a scientific world of extreme uncertainty (or, should I say, a very gray world?).
As a theist who is comfortable accepting the proposals of modern cosmology about the history of the universe—that the sun, moon, stars and this earth came into being via a process—I am inclined to expect that God brought life into existence on this earth in a similar fashion. How it happened in scientific terms remains a deep, deep mystery. An easy way out, it would seem, might be to simply agree with those who just proclaim it to be a miracle and let it go at that. I would prefer an alternative response. The high MA values of biotic molecules may be interpreted as another indication of how extraordinarily life-friendly this Creation is.
Much has been written and argued about the fine-tuning of our universe. Perhaps we could view the existence of living systems that employ low-probability high MA molecules for their existence as an indication that the entire chemical space that gave rise to this phenomenon called life was masterfully intended (designed?) by the Creator to ensure life’s occurrence, though the process leading to it may seem to us to be very gray indeed. This would be fine-tuning of chemistry on a grand scale.
References
1. Smith, Hillary H., et al. “The Grayness of the Origin of Life.” Life 11, 498, 2021.
2. Ibid.
3. Gánti, T. “Biogenesis Itself.” J. Theor. Biol. 187, 583-593, 1997.
4. Smith, et al.
5. Kauffman, Stuart. At Home in the Universe: The Search for the Laws of Self-Organization and Complexity. (New York: Oxford Press, 1995) 31.
6. Ralser, Markus. “An Appeal to Magic? The Discovery of a Non-enzymatic Metabolism and Its Role in the Origins of Life.” Biochemical Journal 475, 2577-2592, 2018.
7. Muchowska, Kamila B., et al. “Nonenzymatic Metabolic Reactions and Life’s Origins.” Chem. Rev. 120, 7708-7744, 2020.
8. Carter, Charles W., Jr. and Wills, Peter R. “The Roots of Genetic Coding in Aminoacyl-tRNA Synthetase Duality.” Annual Review of Biochemistry 90, 349-373, 2021.
9. Carter, Charles W., Jr. “What RNA World? Why a Peptide/RNA Partnership Merits Renewed Experimental Attention.” Life 5, 294-320, 2015.
Larry Funck has a B.S. in Chemistry from Lebanon Valley College, (1964), and a Ph.D. in Inorganic Chemistry from Lehigh University, (1969). He was a Professor of Inorganic Chemistry at Wheaton College (IL), from 1969-2011; a Fulbright Fellow, at the National University of Lesotho, 1995-96. Dr. Funck is currently Emeritus Professor, Wheaton College, and adjunct Professor teaching Origin of Life component of Scientific Theories of Origins course at Wheaton. He is coauthor of Understanding Scientific Theories of Origins, IVP, (2018).
In my opinion the critical juncture between assembly theory and origin of life involves the fundamental question of the probability of life’s occurrence. As Garte suggests in his article, molecules with a higher molecular assembly (MA) index, especially the critically important nucleotides and their polymers, molecules involved in information storage and transmission, would be hard to produce abiotically.
I would suppose that assembly theory proponents would argue that there is an inverse correspondence between a molecule’s MA value and the probability of that molecule occurring in the absence of life. I suspect that they would want to leave open the question of the probability of the origin of life itself.
If the production of high MA molecules needed for life’s origin had to occur in advance of its origin, then it would follow that life’s spontaneous appearance would have an exceedingly low probability. But what if life emerged through a “protracted evolutionary process” that also produced the high MA molecules as part of that process? Then both living systems and high MA molecules associated with life would be the outcome of life’s origin. The question boils down to this: What were the chemical pathways to the production of systems possessing high MA molecules—systems capable of exhibiting life-like behavior, and therefore classifiable as biotic—or, in simpler terms, how did life begin?
Origin-of-life theoretician Stuart Kauffman wrote in 1995 (still true today): “Anyone who tells you that he or she knows how life started on the sere earth… is a fool or a knave” (5).
Origin-of-life theories are abundant and varied. For example, there are the geneticists versus the metabolists. Which had primacy—the production of energy/metabolites or the origin of information coding and reproductive capability? What constituted the first forms of life, and what did they eat—were they heterotrophic, incorporating organic substances, or were they autotrophs, making their own organics from inorganic starting materials in their initial formation and their continued maintenance?
One of the suggested ways that a geochemical to biochemical transition is imagined involves the proposal of catalytic activity by metal atoms/ions on mineral surfaces that initiated the first versions of metabolic processes and led to the development of metalloenzymes. There are plenty of examples of speculative theory along these lines, most extensively in the proposals for life’s origin at deep sea hydrothermal vents.
Perhaps the best experimental support of an origin via autotrophic metabolism comes from recent discoveries of non-enzymatic metabolism. Markus Ralser details in his 2018 review article (6) multiple examples discovered within the last decade of entire metabolic pathways catalyzed by metal ions, particularly Fe(II). He argues that this provides substantial support for the idea that biochemical pathways originated in an abiotic, geochemical context. Ralser suggests that by the use of appropriate analytical techniques, it is likely that many more examples of non-enzymatic metabolic pathways will be discovered.
Muchowska, et al., in a more recent review article (7), discuss the concepts advanced by Ralser and extend the consideration to the possibility of non-enzymatic synthesis pathways for nucleotides that would parallel their biosynthetic pathways. They suggest that the lack of current evidence for such pathways is likely a consequence of the lack of research along those lines. They also argue in favor of a “systems” approach to prebiotic chemistry, which would investigate the possibility of cooperative development of nucleic acids and polypeptides.
So how does this relate to the question of high MA molecules before life’s emergence? In short, the occurrence of complex metabolic pathways driven not by high MA enzymes but by very low MA metal ions suggests the possibility that the high MA molecules may have emerged in the process of the gradual progression towards systems that would have increasingly biotic characteristics and be eventually characterized as alive.
Arguably the most critical step in the progression towards life involved the appearance of the genetic coding system and accompanying enzymatic apparatus. There are several theories about how this occurred, but none has gained wide acceptance to date. In my opinion, the most attractive one has been advanced by Carter and Wills (8). Their theory, which is empirically based, rests principally on a proposal of intimate cooperation between RNA and peptides in the emergence of the code through the formation of “complexes involving structurally complementary oligonucleotides and dipeptides” (9). These complexes are proposed to have originated long before genuine code expression occurred involving simple versions of tRNA and aaRS proteins.
My guess is that were Carter or Wills to be asked about the existence of these relatively high MA value molecular complexes well in advance of a functional coding system, they would probably reply in a manner similar to that proposed above—the high MA oligonucleotide/dipeptide complexes were merely intermediates on the path to life in the “protracted evolutionary process.”
So what should we conclude? Should the high MA values for biomolecules be viewed as problematic for the origin-of-life theoretician? I would suggest they will be troublesome if the theoretician is uncomfortable living in a scientific world of extreme uncertainty (or, should I say, a very gray world?).
As a theist who is comfortable accepting the proposals of modern cosmology about the history of the universe—that the sun, moon, stars and this earth came into being via a process—I am inclined to expect that God brought life into existence on this earth in a similar fashion. How it happened in scientific terms remains a deep, deep mystery. An easy way out, it would seem, might be to simply agree with those who just proclaim it to be a miracle and let it go at that. I would prefer an alternative response. The high MA values of biotic molecules may be interpreted as another indication of how extraordinarily life-friendly this Creation is.
Much has been written and argued about the fine-tuning of our universe. Perhaps we could view the existence of living systems that employ low-probability high MA molecules for their existence as an indication that the entire chemical space that gave rise to this phenomenon called life was masterfully intended (designed?) by the Creator to ensure life’s occurrence, though the process leading to it may seem to us to be very gray indeed. This would be fine-tuning of chemistry on a grand scale.
References
1. Smith, Hillary H., et al. “The Grayness of the Origin of Life.” Life 11, 498, 2021.
2. Ibid.
3. Gánti, T. “Biogenesis Itself.” J. Theor. Biol. 187, 583-593, 1997.
4. Smith, et al.
5. Kauffman, Stuart. At Home in the Universe: The Search for the Laws of Self-Organization and Complexity. (New York: Oxford Press, 1995) 31.
6. Ralser, Markus. “An Appeal to Magic? The Discovery of a Non-enzymatic Metabolism and Its Role in the Origins of Life.” Biochemical Journal 475, 2577-2592, 2018.
7. Muchowska, Kamila B., et al. “Nonenzymatic Metabolic Reactions and Life’s Origins.” Chem. Rev. 120, 7708-7744, 2020.
8. Carter, Charles W., Jr. and Wills, Peter R. “The Roots of Genetic Coding in Aminoacyl-tRNA Synthetase Duality.” Annual Review of Biochemistry 90, 349-373, 2021.
9. Carter, Charles W., Jr. “What RNA World? Why a Peptide/RNA Partnership Merits Renewed Experimental Attention.” Life 5, 294-320, 2015.
Larry Funck has a B.S. in Chemistry from Lebanon Valley College, (1964), and a Ph.D. in Inorganic Chemistry from Lehigh University, (1969). He was a Professor of Inorganic Chemistry at Wheaton College (IL), from 1969-2011; a Fulbright Fellow, at the National University of Lesotho, 1995-96. Dr. Funck is currently Emeritus Professor, Wheaton College, and adjunct Professor teaching Origin of Life component of Scientific Theories of Origins course at Wheaton. He is coauthor of Understanding Scientific Theories of Origins, IVP, (2018).